
Breve descripción del origen, formulación y crítica de la TPV, agosto de 2021
El IPV se complace en publicar el siguiente comentario del Dr. Stephen Porges sobre su Teoría Polivagal (TPV), que incluye una descripción introductoria, información previa necesaria y los principios primarios. La última sección aborda las críticas a la teoría desde su publicación original en 1995.
Índice
- Introducción a la TPV
- Información previa para comprender la TPV
- Principios básicos de la TPV
- Principio 1: El estado autónomo funciona como una «variable interviniente».
- Principio 2: Las vías vagales ventrales funcionan como un freno vagal transitorio que favorece la atención, la vigilancia, el esfuerzo mental, la preparación para el movimiento y la anticipación de amenazas.
- Principio 3: La reactividad autónoma está organizada en una secuencia de respuesta evolutiva caracterizada por la inhibición jerárquica y la disolución
- Principio 4: La neurocepción es el proceso implicado en la detección de señales de amenaza o seguridad y el ajuste del estado autónomo para apoyar estrategias de supervivencia adaptativas.
- Principio 5: La teoría polivagal hace hincapié en las diferencias neuroanatómicas y funcionales únicas que evolucionaron en los mamíferos en relación con otros vertebrados y que conducen a nuestra herencia social como mamíferos.
- Principio 6: Las críticas tergiversan la teoría polivagal
Introducción
Desde la publicación de la teoría polivagal (PVT) en 1995, el Dr. Porges ha publicado actualizaciones ocasionales que abordan su formulación, siendo la teoría polivagal una teoría con base científica que proporciona explicaciones sobre cómo nuestro estado autónomo influye y es influido por los retos dinámicos de la vida. La teoría, como perspectiva, incide en varios aspectos de la experiencia humana, así como en las experiencias de otros mamíferos que comparten características de la regulación neuronal del sistema nervioso autónomo. Se trata de una teoría transdisciplinar que extrae temas comunes a través de un amplio abanico de disciplinas.
La teoría polivagal fue presentada por el neurocientífico del comportamiento, Stephen W. Porges, en su discurso presidencial ante la Sociedad de Investigación Psicofisiológica en Atlanta (Georgia) el 8 de octubre de 1994. La charla se publicó posteriormente en Psychophysiology (1995) con el título Orientarse en un mundo defensivo: Modificaciones en los mamíferos de nuestra herencia evolutiva. Una teoría polivagal (Porges, 1995).
El título representa una sinopsis sucinta de la teoría, en la que se analiza la transición filogenética desde los reptiles asociales hasta los mamíferos sociales haciendo hincapié en los cambios convergentes en la regulación neuronal del sistema nervioso autónomo y las consecuencias bioconductuales adaptativas.
Desde el punto de vista funcional, a diferencia de sus antepasados reptiles, los mamíferos tienen una rama más reciente del vago que ejerce una influencia cardioinhibidora sobre el corazón y que emerge de una zona ventral del tronco encefálico (el núcleo ambiguo).
En cambio, otras clases de vertebrados (por ejemplo, reptiles, anfibios, etc.) tenían vías cardioinhibitorias vagales que emergían sólo de una zona dorsal del tronco encefálico (es decir, el núcleo motor dorsal del vago). Así, la teoría llamó explícitamente la atención sobre la función adaptativa de la vía vagal ventral en la definición de las características de los mamíferos sociales, incluida la regulación mutua de las reacciones de amenaza, del estado fisiológico de calma y la socialidad.
La versión publicada de la charla documenta las conclusiones citando una extensa lista de referencias de más de 100 publicaciones científicas de varias disciplinas. Publicaciones posteriores ampliaron y refinaron la teoría (Porges, 1998,2001,2003, 2007, 2021 y Porges y Lewis, 2010) y validaron la metodología utilizada para monitorizar la vía cardioinhibitoria ventral vagal (Lewis y col. 2012).
La teoría ha ganado terreno en diversas disciplinas y ha sido citada en miles de artículos revisados por pares (véase Google Scholar). La investigación fundacional que desarrolló la teoría contó con 38 años de financiación continua (1975-2013) del Instituto Nacional de Salud de EEUU. Desde su presentación, la teoría ha sido citada en más de 10.000 artículos revisados por pares (véase Google Scholar).
La teoría polivagal recibe su nombre del vago, un nervio craneal que forma el componente primario del sistema nervioso autónomo conocido como sistema nervioso parasimpático. La visión tradicional del vago es que es el nervio que apoya la salud, el crecimiento y la restauración o, simplemente, los procesos homeostáticos. En la visión tradicional, el sistema nervioso parasimpático tiene la capacidad de compensar y neutralizar las costosas demandas metabólicas del sistema nervioso simpático que apoya los comportamientos de lucha y huida. Esta visión es sólo parcialmente correcta porque el sistema nervioso parasimpático también desempeña un papel en las estrategias defensivas de supervivencia.
La teoría es una declaración y aporta una solución a la paradoja vagal observada en neonatología. La paradoja se formuló contrastando observaciones bien documentadas de influencias vagales tanto protectoras como letales. Específicamente, un supuesto fenómeno vagal, la bradicardia clínica (disminución masiva de la frecuencia cardiaca), es potencialmente letal en los recién nacidos prematuros de alto riesgo y durante partos complicados, mientras que la arritmia sinusal respiratoria, también un supuesto fenómeno vagal, es mayor en los recién nacidos sanos que en los prematuros de alto riesgo (véase Porges, 1992).
La teoría resolvió la paradoja proponiendo que la bradicardia y la arritmia sinusal respiratoria eran salidas funcionales de dos áreas diferentes del tronco encefálico que contenían vías cardioinhibitorias que tenían reflejos diferentes en el patrón de la frecuencia cardiaca. El artículo fundacional documentaba que los mamíferos modernos tenían un «oscilador cardiorrespiratorio común» (véase Richter y Spyer, 1990) en el que sólo participaba la vía vagal ventral, mientras que la vía vagal cardioinhibitoria dorsal sostenía la bradicardia.
Al igual que en los antepasados reptiles, la bradicardia masiva era un componente de una reacción ante una amenaza que a menudo se observa como muerte fingida en pequeños mamíferos. Investigaciones posteriores (Reed y col., 1999) documentaron que la monitorización del estado de la vía vagal ventral mediante la cuantificación de la arritmia sinusal respiratoria identificaba la vulnerabilidad a la bradicardia clínica en humanos durante el proceso del parto.
Desde el punto de vista funcional, la teoría polivagal proporciona una base para una ciencia cerebro-cuerpo integrada y expansiva que hace hincapié en la comunicación bidireccional entre los órganos viscerales y las estructuras cerebrales a través de las vías vagales.
La teoría se basa en la investigación que rompió paradigmas de varios científicos visionarios, que introdujeron conceptos que se han reformulado dentro de la teoría polivagal, como el medio interno (Claude Bernard), la homeostasis (Walter Cannon), la evolución (Charles Darwin), la disolución (John Hughlings Jackson), la activación (Robert Yerkes y John Dodson), la lucha/huida (Walter Cannon) y un sistema nervioso unificado que integra las funciones cerebrales y corporales (Walter Hess). Al extraer y combinar principios aceptados descubiertos por estos científicos fundacionales, la teoría polivagal proporciona un nuevo conocimiento de la regulación neuronal del sistema nervioso autónomo que hace hincapié en la comunicación bidireccional entre el cerebro y los órganos viscerales.
Respetando la literatura científica histórica, la teoría actualiza la conceptualización del sistema nervioso autónomo mediante un modelo transdisciplinar que extrae principios básicos que han surgido de forma independiente y han sido documentados en disciplinas dispares. La teoría polivagal, como teoría, conduce a hipótesis plausibles que tienen amplias aplicaciones en los ámbitos dispares de la educación, el desarrollo infantil, la salud mental, la salud física, los entornos laborales y las instituciones sociales, además de proporcionar conocimientos sobre el bienestar de otros mamíferos sociales.
Información previa para comprender la teoría polivagal
1. La relación entre el estado autónomo y las conductas defensivas
En la mayoría de los vertebrados, los dos principales sistemas de defensa son la lucha o la huida y la inmovilización.
- Los comportamientos de lucha o huida permiten al organismo huir o defenderse cuando se ve amenazado. Estos comportamientos requieren la rápida accesibilidad de los recursos a movilizar a través de la activación del sistema nervioso simpático, metabólicamente costoso.
- La inmovilización es un sistema de defensa más antiguo, que comparten prácticamente todos los vertebrados. En contraste con la estrategia de movilización metabólicamente costosa, la inmovilización es un intento adaptativo de reducir las exigencias metabólicas (por ejemplo, posibilidades reducidas de alimento y oxígeno) y de parecer inanimado (por ejemplo, fingir la muerte).
En contraste con la rápida activación del sistema nervioso simpático necesaria para promover los comportamientos de lucha o huida, los comportamientos defensivos de inmovilización requieren un apagado masivo de la función autónoma a través de una vía vagal dentro del sistema nervioso parasimpático.
Antes de la teoría polivagal, este último sistema de defensa se ignoraba o minimizaba, mientras que se suponía que la amenaza provocaba únicamente una reacción de lucha-huida movilizada que requería la excitación simpática. La dificultad para conceptualizar una reacción de defensa vagal se debía, en parte, a la bien documentada retirada del tono vagal cardioinhibitorio durante la amenaza que optimiza la expresión de la activación simpática.
Así, se asumió que un tono vagal bajo era un marcador de una reacción de amenaza. Sin una conceptualización de las dos vías vagales, la explicación neurofisiológica de la bradicardia clínica estaba incompleta. La teoría polivagal replanteó la investigación de la base autónoma de las reacciones de amenaza. Parsimoniosamente, la retirada del tono vagal ventral durante la amenaza eliminaba la competencia por la activación simpática eficiente para aumentar el gasto cardíaco con el fin de apoyar la movilización y aumentaba la vulnerabilidad a un síncope masivo inducido por la vía vagal dorsal que apoyaría la inmovilización.
Esta secuencia se observa durante partos humanos difíciles en los que la bradicardia potencialmente mortal va precedida de una disminución de la variabilidad de la frecuencia cardiaca y taquicardia (véase Reed y col.).
2. El estudio filogenético de los sistemas nerviosos autónomos de los vertebrados identifica tres circuitos primarios
A través de los procesos evolutivos de la transición filogenética desde los reptiles asociales hasta los mamíferos sociales, evolucionó una segunda vía vagal cardioinhibitoria que tenía la capacidad de regular a la baja ambas formas de defensa.
Esta segunda vía vagal se observa en mamíferos y no en reptiles. Además, a diferencia de los reptiles y otros vertebrados con vías vagales cardioinhibitorias, las estructuras anatómicas que regulan este componente mamífero del vago interactúan en el tronco encefálico con estructuras que regulan los músculos estriados de la cara y la cabeza para proporcionar un sistema integrado de conexión social (véase más abajo).
A través de estudios de la literatura de neuroanatomía comparada se pueden estimar las transiciones filogenéticas en la regulación neuronal del sistema nervioso autónomo de los vertebrados. Las conclusiones plausibles apoyan la identificación de tres circuitos que siguen una secuencia filogenética en la que el circuito neuronal más antiguo emerge del núcleo dorsal del vago, seguido del sistema simpático espinal, y el más evolucionado emerge del núcleo ventral del vago.
Durante la transición a la tercera etapa, cuando evolucionaron las vías vagales ventrales, las células de origen del vago migraron del núcleo dorsal del vago al núcleo ambiguo. Esta transición destaca por la mielinización de las fibras motrices vagales cardioinhibitorias ventrales y su integración en el tronco encefálico para regular una familia de vías motrices (es decir, las vías eferentes viscerales especiales) que controlan los músculos estriados de la cara y la cabeza.
Aunque los circuitos antiguos se han reutilizado y potencialmente modificado a través de procesos evolutivos, las características se han transmitido a los mamíferos sociales, incluidos los humanos. Un apoyo adicional a la secuencia filogenética procede de la literatura sobre el desarrollo, que extrae tendencias madurativas de estudios de autopsias de embriones, fetos y mamíferos jóvenes después del parto, incluidos datos humanos (véase Porges y Furman, 2011).
3. La aparición del sistema de conexión social: Perspectivas de la evolución y el desarrollo
La integración de las vías vagales cardiacas mielinizadas con la regulación neuronal de la cara y la cabeza dio lugar al sistema de conexión social de los mamíferos. Como se ilustra en la figura siguiente, las salidas del sistema de conexión social consisten en vías motrices que regulan los músculos estriados de la cara y la cabeza (es decir, somatomotrices) y los músculos lisos y cardíacos del corazón y los bronquios (es decir, visceromotrices).
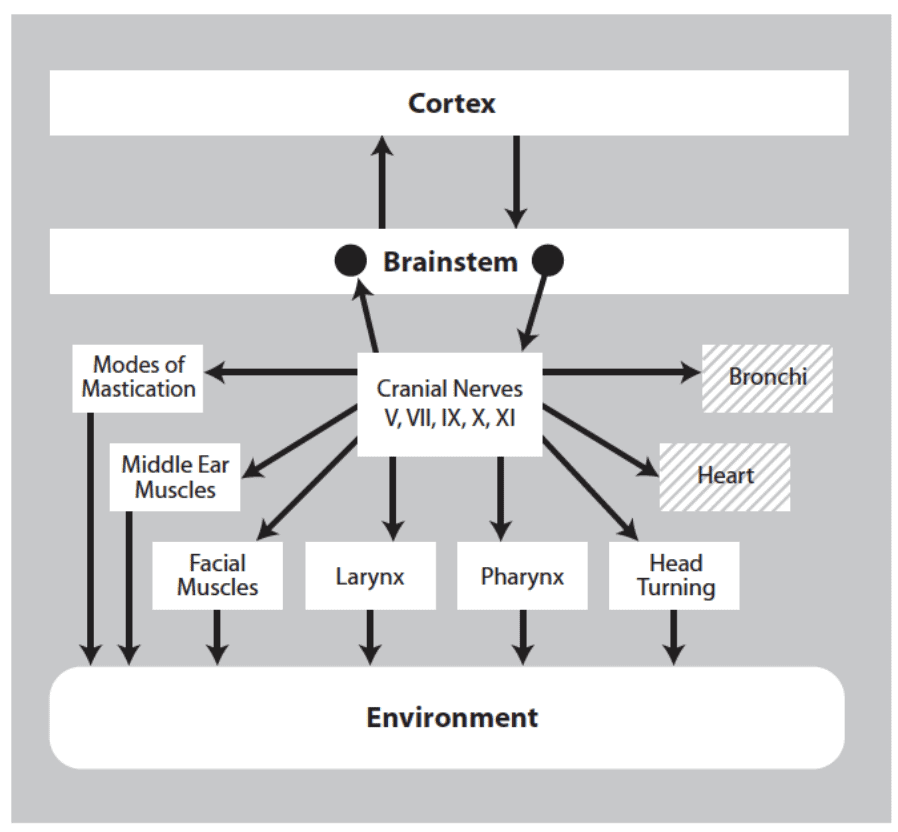
En el componente somatomotor intervienen vías eferentes viscerales especiales que regulan los músculos estriados de la cara y la cabeza. En el componente visceromotor intervienen las vías vagales mielinizadas supradiafragmáticas (por encima del diafragma) que regulan el corazón y los bronquios.
Funcionalmente, el sistema de conexión social surge de una conexión cara-corazón que coordina el corazón con los músculos de la cara y la cabeza. La función inicial del sistema es coordinar la succión, la deglución, la respiración y la vocalización. Una coordinación atípica de este sistema en los primeros años de vida es un indicador de dificultades posteriores en el comportamiento social y la regulación emocional.
Cuando está plenamente desarrollado, se manifiestan dos importantes características bioconductuales de este sistema:
- En primer lugar, el estado corporal se regula de manera eficiente para promover el crecimiento y la restauración (por ejemplo, la homeostasis visceral). Funcionalmente, esto se consigue mediante un aumento de la influencia de las vías motrices vagales mielinizadas sobre el marcapasos cardíaco para ralentizar el ritmo cardíaco, inhibir los mecanismos de lucha o huida del sistema nervioso simpático, amortiguar el sistema de respuesta al estrés del eje hipotalámico-hipofisario-suprarrenal (HHS) (responsable de la liberación de cortisol) y reducir la inflamación modulando las reacciones inmunitarias (por ejemplo, las citocinas; para una revisión, véase Porges, 2007).
- En segundo lugar, la conexión rostro-corazón de los mamíferos filogenéticamente funciona para transmitir el estado fisiológico a través de la expresión facial y la prosodia (entonación de la voz), así como regular los músculos del oído medio para optimizar la escucha específica de cada especie dentro de la banda de frecuencia utilizada para la comunicación social (Kolacz, Lewis y Porges,2018; Porges, 2007, 2009, 2011; Porges y Lewis, 2010 ).
Este sistema emergente de conexión social proporciona el mecanismo para la corregulación del estado fisiológico, ya que los mamíferos transmiten señales de seguridad y peligro -a través de vocalizaciones, gestos con la cabeza y expresiones faciales- a sus conespecíficos. El sistema de conexión social permitió a los mamíferos cooptar algunas de las características de los sistemas de defensa de los vertebrados para promover interacciones sociales como el juego y la intimidad.
Estos cambios en el sistema nervioso autónomo dotaron a los mamíferos de mecanismos neuronales para promover los estados bioconductuales necesarios para el cuidado de las crías, la reproducción y el comportamiento cooperativo. En cambio, los efectos psicológicos y conductuales adversos de la amenaza parecen dirigirse a una alteración del sistema de conexión social, su gestión de las reacciones de defensa y su contribución a la corregulación y las conductas cooperativas, como la intimidad y el juego.
Las vías motrices vagales ventrales mielinizadas «más nuevas» regulan los órganos supradiafragmáticos (por ejemplo, el corazón y los pulmones) y se integran en el tronco encefálico con estructuras que regulan los músculos estriados de la cara y la cabeza a través de vías eferentes viscerales especiales, lo que da lugar a un sistema funcional de conexión social.
Este circuito vagal más reciente ralentiza el ritmo cardíaco y favorece los estados de calma necesarios para las interacciones sociales. El circuito vagal ventral acoplado con otros circuitos autónomos apoya el juego social (es decir, el vagal ventral acoplado con la activación simpática) y la intimidad segura (es decir, el vagal ventral acoplado con el circuito vagal dorsal). Por lo tanto, el vago de los mamíferos tiene propiedades que promueven estados que contienen el rango de respuesta de todos los componentes del sistema nervioso autónomo y funcionalmente impide que el sistema pase a estados de defensa.
La tabla siguiente proporciona un mapa plausible de los perfiles autónomos asociados a diferentes categorías de comportamiento. La tabla hace hincapié en el papel coordinador del circuito vagal ventral en la expansión de experiencias positivas y sociales mediante la integración de todos los atributos neuronales implicados en la regulación de la función autónoma.
| Comportamiento emergente | Dorsal Vagal | Simpático | Ventral Vagal |
| Tranquilo, socialmente corregulado | X | ||
| Juego social y baile | X | X | |
| Agresivo, irritable, combativo, lucha-huida, estrés crónico, dolor | X | ||
| Paralización | X | X | |
| Calma, intimidad compartida | X | X | |
| Desconexión, muerte fingida | X |
Principios básicos de la teoría polivagal
Principio 1: El estado autónomo funciona como una «variable interviniente».
La teoría polivagal propone que el estado fisiológico es una parte fundamental, y no un correlato, de la emoción o el estado de ánimo. Según esta teoría, el estado autónomo funciona como una variable interviniente que sesga nuestra detección y evaluación de las señales del entorno. Dependiendo del estado fisiológico, las mismas señales se evaluarán de forma refleja como neutras, positivas o amenazadoras (véase la neurocepción más adelante).
Desde el punto de vista funcional, un cambio de estado cambiará el acceso a diferentes estructuras del cerebro y favorecerá la comunicación social o los comportamientos defensivos de lucha o huida o de desconexión.
La investigación contemporánea sobre el impacto de la estimulación del nervio vago en la función cognitiva y la regulación de las emociones respalda este modelo (Groves y Brown, 2005).
La teoría hace hincapié en un vínculo bidireccional entre cerebro y vísceras, que explicaría cómo los pensamientos cambian la fisiología y el estado fisiológico influye en los pensamientos. Cuando una persona cambia sus expresiones faciales, la entonación de su voz, el patrón de su respiración y su postura, también cambia su fisiología a través de circuitos en los que intervienen las vías vagales mielinizadas que van al corazón.
La teoría hace hincapié en que el estado fisiológico influye en la actividad conductual y mental, incluidas la función cognitiva y la accesibilidad al aprendizaje, así como la emocionalidad y la socialidad. También subraya que el estado fisiológico puede influir potencialmente en los resultados de salud y los tratamientos médicos.
El modelo subraya que el estado autónomo funciona como una variable «interviniente» que amortigua o exacerba el impacto perturbador de los cambios en los procesos homeostáticos. En general, el estado autónomo puede conceptualizarse como un apoyo a las funciones homeostáticas de salud, crecimiento o restauración o, alternativamente, como una alteración de los procesos homeostáticos para apoyar una reacción de amenaza generalizada.
Desde el punto de vista funcional, el estado fisiológico se conceptualiza como una «plataforma neuronal» de la que dependen los procesos conductuales, mentales, sociales y fisiológicos. Esto conduce a hipótesis plausibles y comprobables de que los cambios en la «plataforma neuronal» optimizarán o amortiguarán el proceso dependiente.
Existe una amplia bibliografía que documenta experimentos en los que las diferencias individuales, evolutivas y estatales en la función autónoma, a menudo medidas por la variabilidad de la frecuencia cardiaca como índice de regulación vagal, están relacionadas con respuestas cognitivas (toma de decisiones), autónomas (reactividad de la frecuencia cardiaca) y físicas (tiempo de reacción).
En las investigaciones que evalúan el efecto de las experiencias infantiles adversas en el comportamiento adulto, los resultados están mediados de forma fiable por un sistema nervioso autónomo resintonizado que conceptualmente puede caracterizarse como «bloqueado» en un estado de amenaza crónica. Estos estudios, al documentar que muchas relaciones «estímulo-respuesta» están mediadas por el estado autónomo, apoyan la teoría al proporcionar pruebas que respaldan el papel del estado autónomo como variable interviniente.
Este principio conduce a una reconceptualización y redefinición del «estrés«. En lugar de la visión predominante del estrés, que a menudo da lugar a una circularidad entre el acontecimiento estresante y la reacción de estrés, la teoría polivagal proporciona una definición funcional objetiva. Desde el punto de vista de la teoría polivagal, el estrés es simplemente una alteración crónica de la función homeostática.
Las alteraciones transitorias del sistema nervioso autónomo que dan lugar a una rápida recuperación no son estrés, ni tampoco las consecuencias de la activación simpático-suprarrenal necesaria para aumentar la actividad metabólica asociada al movimiento. A corto plazo, el sistema nervioso autónomo se ajusta sin problemas para soportar las demandas metabólicas y, en el caso óptimo, se recupera rápidamente en un estado que soporta las funciones homeostáticas.
En lugar de depender del ambiguo concepto de estrés, la teoría polivagal sugiere que el perfil autónomo se interprete desde una perspectiva adaptativa, preguntando si el sistema nervioso autónomo se encuentra en un estado que favorece las reacciones de amenaza o las funciones homeostáticas de calma.
Las reacciones de amenaza, especialmente un sistema nervioso autónomo bloqueado en un estado de amenaza, se identifica fácilmente por tener un tono cardioinhibitorio vagal ventral bajo y un umbral bajo para ser simpáticamente reactivo y potencialmente vulnerable a la desconexión vagal dorsal. Este sistema nervioso autónomo resintonizado está bien preparado para actuar a la defensiva, lo que incluye apoyar la hipersensibilidad y la hipervigilancia, al tiempo que compromete la regulación neuronal de los órganos viscerales.
Principio 2: Las vías vagales ventrales funcionan como un freno vagal transitorio que favorece la atención, la vigilancia, el esfuerzo mental, la preparación para el movimiento y la anticipación de amenazas.
La distinción entre alteración transitoria y crónica de los procesos homeostáticos condujo a la conceptualización del freno vagal (véase Porges, 1996). Desde finales de la década de 1960 (por ejemplo, Porges, 1969, 1972 y Porges y Walter, 1976), los cambios en la variabilidad de la frecuencia cardiaca se utilizaron como indicadores de la atención sostenida y el esfuerzo mental.
Además, cientos de estudios en docenas de laboratorios documentaron que los cambios en una variedad de índices de variabilidad de la frecuencia cardiaca, incluida la arritmia sinusal respiratoria, se deprimían durante formas de atención, anticipación y durante demandas metabólicas como el ejercicio (Hatfield y col.). La teoría polivagal atribuyó a la vía vagal ventral el mecanismo subyacente al freno vagal.
Investigaciones posteriores han destacado el papel de la recuperación de la arritmia sinusal respiratoria (Calkins…, Dale…) como un índice importante del comportamiento social y la regulación del estado, lo que lleva a inferir que la vía cardioinhibitoria ventral vagal está implicada tanto en la autorregulación como en la corregulación y su regulación subyace al concepto de resiliencia.
Una vez introducida la conceptualización del freno vagal dentro de la teoría polivagal (Porges, 1996), los cambios comúnmente observados en las medidas globales de la variabilidad de la frecuencia cardíaca y el componente «vagal» más específico de la arritmia sinusal respiratoria durante los desafíos psicológicos y físicos pudieron entenderse desde una perspectiva neurobiológica. Esta perspectiva ha adoptado la tecnología para cuantificar señales neuronales específicas como la arritmia sinusal respiratoria como índice preciso del tono vagal ventral (Lewis y col., 2012).
Principio 3: La reactividad autónoma está organizada en una secuencia de respuesta evolutiva caracterizada por la inhibición jerárquica y la disolución.
La teoría hace hincapié en que, en respuesta a los desafíos, el estado autónomo cambia a través de una secuencia definida por la evolución a la inversa o la disolución. La disolución es un concepto introducido por John Hughlings Jackson que hace hincapié en la inhibición jerárquica de circuitos neuronales evolutivamente más antiguos por circuitos más nuevos. El sistema nervioso humano, al igual que el de otros mamíferos, evolucionó no sólo para sobrevivir en entornos seguros, sino también para promover la supervivencia en contextos peligrosos y potencialmente mortales.
Para lograr esta flexibilidad adaptativa, el sistema nervioso autónomo de los mamíferos, además de la vía vagal mielinizada que se integra en el sistema de conexión social, conservó dos circuitos neuronales más primitivos para regular las estrategias defensivas (es decir, los comportamientos de lucha o huida y de amenaza de muerte). Es importante señalar que el comportamiento social, la comunicación social y la homeostasis visceral son incompatibles con los estados neurofisiológicos que sustentan la defensa.
Las estrategias de respuesta polivagal a los desafíos están ordenadas filogenéticamente, siendo los componentes más nuevos del sistema nervioso autónomo los que responden primero. Este modelo de reactividad autónoma es coherente con el concepto de disolución de John Hughlings Jackson, en el que proponía que «las estructuras nerviosas superiores inhiben (o controlan) las inferiores y, por lo tanto, cuando las superiores quedan repentinamente sin función, las inferiores aumentan su actividad» (1882, p. 412). En esta jerarquía de respuestas adaptativas, primero se utiliza el circuito de conexión social más reciente; si ese circuito no proporciona seguridad, se recurre secuencialmente a los circuitos más antiguos.
Principio 4: La neurocepción es el proceso implicado en la detección de señales de amenaza o seguridad y el ajuste del estado autónomo para apoyar estrategias adaptativas de supervivencia.
La teoría polivagal propone que la evaluación neuronal del riesgo no requiere una percepción consciente y funciona a través de circuitos neuronales que compartimos con nuestros ancestros vertebrados filogenéticos. Así, se introdujo el término neurocepción para enfatizar un proceso neuronal, distinto de la percepción, capaz de detectar características ambientales y viscerales que son seguras, peligrosas o amenazan la vida (Porges, 2003, 2004). La neurocepción es un mecanismo reflexivo capaz de cambiar instantáneamente el estado fisiológico.
La neurocepción implica mecanismos descendentes y ascendentes. En los seres humanos, se supone que el proceso se inicia a través de vías descendentes en las que intervienen áreas corticales situadas en el córtex temporal o cerca de él, que interpretan de forma refleja las señales de amenaza y seguridad.
Estas zonas del córtex son sensibles a la intencionalidad de los movimientos biológicos, como voces, caras, gestos y movimientos de las manos. La neurocepción es la capacidad del sistema nervioso para reaccionar ante la intención de estos movimientos. La neurocepción descodifica e interpreta funcionalmente el supuesto objetivo de los movimientos y sonidos de objetos inanimados y vivos.
Así, la neurocepción de personas conocidas y de individuos con voces adecuadamente prosódicas y rostros cálidos y expresivos se traduce con frecuencia en una interacción social positiva, que fomenta la sensación de seguridad. El estado autónomo responde a la detección descendente de riesgo o seguridad. La reacción autónoma envía información sensorial sobre las sensaciones corporales al cerebro, donde se interpretan y se sienten conscientemente. La rama ascendente de la neurocepción equivale funcionalmente a la interocepción.
Así pues, aunque a menudo no somos conscientes de los estímulos que desencadenan las distintas respuestas de la neurocepción, en general somos conscientes de las reacciones de nuestro cuerpo (es decir, las sensaciones viscerales) plasmadas en otras manifestaciones autónomas que sustentan comportamientos adaptativos (es decir, conexión social, lucha/huida, desconexión).
Se puede encontrar una forma de neurocepción en prácticamente todos los organismos vivos, independientemente del desarrollo del sistema nervioso. De hecho, podría afirmarse que los organismos unicelulares e incluso las plantas tienen un sistema nervioso primordial que responde a la amenaza.
Como mamíferos, estamos familiarizados con las reacciones al dolor (es decir, la nocicepción), un tipo de neurocepción. Reaccionamos al dolor antes de ser capaces de identificar la fuente del estímulo o incluso de ser conscientes de la lesión. La detección de la amenaza parece ser común a todas las especies de vertebrados. Sin embargo, los mamíferos tienen una capacidad de neurocepción ampliada en la que no sólo reaccionan instantáneamente a la amenaza, sino que también responden instantáneamente a las señales de seguridad.
Es esta última característica la que permite a los mamíferos regular a la baja las estrategias defensivas para fomentar la socialidad, permitiendo la proximidad psicológica y física sin las consecuencias de las lesiones. Es este mecanismo tranquilizador el que ajusta de forma adaptativa la regulación central de la función autónoma para amortiguar la activación simpática y proteger el sistema nervioso central dependiente del oxígeno, especialmente la corteza, de las reacciones metabólicamente conservadoras del complejo vagal dorsal (por ejemplo, desmayos, muerte fingida).
Aprovechar el poder de la neurocepción para regular a la baja un estado autónomo que apoyaría las estrategias defensivas es una característica primordial de las terapias basadas en la teoría polivagal. Por ejemplo, el protocolo Safe and Sound, la intervención acústica desarrollada por Porges, aplica un algoritmo informático para amplificar las características prosódicas de las vocalizaciones humanas que funcionan como un estimulador acústico del nervio vago (página web). El protocolo Safe and Sound se utiliza frecuentemente con otras estrategias terapéuticas para mejorar la accesibilidad a la terapia calmando el sistema nervioso autónomo.
Principio 5: La teoría polivagal hace hincapié en las diferencias neuroanatómicas y funcionales únicas que evolucionaron en los mamíferos en relación con otros vertebrados y que conducen a nuestra herencia social como mamíferos.
La teoría polivagal se centra en las diferencias, más que en las similitudes, entre las especies de vertebrados y, especialmente, en los cambios neuroanatómicos y funcionales observados durante la transición desde los reptiles asociales hasta los mamíferos sociales.
Por ejemplo, existen distinciones neuroanatómicas y funcionales entre la arritmia del seno respiratorio de los mamíferos y las interacciones entre la frecuencia cardiaca y respiratoria en otros vertebrados. Pero sólo los mamíferos tienen un oscilador respiratorio central común bien definido que envía un ritmo respiratorio desde el tronco encefálico tanto al corazón como a los bronquios.
Esta información fluye a través de las neuronas vagales que se originan en el núcleo ambiguo. De hecho, el oscilador puede conceptualizarse como una propiedad emergente de las interacciones entre las estructuras reguladas por el núcleo ambiguo, incluidas la laringe y la faringe (Richter y Spyer, 1990).
Esto, por supuesto, no es coherente con las características de un núcleo ambiguo primitivo o del núcleo dorsal del vago que podrían observarse en los vertebrados que precedieron a los mamíferos. La observación de un oscilador central común es funcionalmente exclusiva de los mamíferos y es la base neurofisiológica que permite la cuantificación de la arritmia del seno respiratorio para que funcione como un portal para obtener una medición sensible y precisa del tono cardiaco vagal ventral.
La teoría utiliza la evolución para extraer una secuencia filogenética de la regulación autónoma. Esta secuencia identifica etapas durante la evolución de los vertebrados en las que surgieron un sistema nervioso simpático espinal y las dos vías vagales, que se hicieron funcionales mediante la maduración en los mamíferos. Sería difícil argumentar que la secuencia no se produce, aunque sería posible identificar similitudes antecedentes en la mayoría de los vertebrados, independientemente de su clase o grupo.
La cuestión no es si existen similitudes en los vertebrados ancestrales, sino más bien cómo se han adaptado estos circuitos para proporcionar un sistema nervioso autónomo único en los mamíferos que está íntimamente entrelazado con el comportamiento social corregulador.
La evolución transformó los atributos del sistema nervioso autónomo en un sistema integrado de conexión social que incorporaba un área de comunicación del tronco encefálico (es decir, el complejo vagal ventral) que regulaba, a través de vías eferentes viscerales especiales, los músculos estriados de la cara y la cabeza y coordinaba estos procesos con la regulación vagal del corazón y los bronquios.
En los mamíferos, el complejo vagal ventral permite coordinar un sistema de succión, deglución, respiración y vocalización con la regulación vagal del corazón. A medida que la neuroanatomía de este circuito digestivo madura, el circuito se convierte en un sistema funcional de conexión social (véase más arriba) que permite comunicar el estado fisiológico a los conespecíficos mediante la expresión facial y las vocalizaciones.
En los mamíferos, estas estructuras y su regulación neuronal se han modificado a través de la evolución para apoyar funciones únicas para la supervivencia de los mamíferos, como la lactancia y la comunicación social. Esta transición en la neuroanatomía y la función sienta las bases para comprender que para los humanos, al igual que para otros mamíferos, la conexión y las relaciones de confianza son expresiones directas de nuestro imperativo biológico y están integradas en nuestra biología.
La teoría polivagal depende de los procesos que los teóricos de la evolución describen como exadaptación y cooptación. Estos procesos implican modificaciones que cambian la función de una estructura durante la evolución. Por ejemplo, una estructura puede evolucionar porque cumplía una función, pero posteriormente puede pasar a cumplir otra. La exadaptación y la cooptación son estrategias comunes de reutilización de estructuras vestigiales tanto en la anatomía como en el comportamiento.
La teoría polivagal es agnóstica sobre las presiones evolutivas que dan lugar a la selección de cambios específicos. En cambio, la teoría es más descriptiva «filogenéticamente» y se centra más en el resultado funcional de la reutilización. Más concretamente, la teoría se interesa por cómo las estructuras reguladas por el complejo vagal ventral fueron reutilizadas para regular un sistema integrado de conexión social que proporciona el portal principal para comprometerse socialmente y comunicarse, ingerir y calmarse.
Los conocimientos neuroanatómicos actuales documentan el refinamiento del complejo vagal ventral en los mamíferos. Por lo tanto, aunque el vago ventral (es decir, el que se origina en el núcleo ambiguo) puede tener su origen en los reptiles (Taylor, 2014), parece que sólo en los mamíferos esta vía se ha reconvertido para transmitir y responder a las señales sociales, a través de la neurocepción, como un potente mediador del estado autónomo.
Principio 6: Las críticas tergiversan la teoría polivagal
Las críticas a la teoría polivagal no reconocen el énfasis de la teoría en los cambios estructurales y funcionales únicos en la regulación del sistema nervioso autónomo que se produjeron durante la transición desde reptiles asociales hasta mamíferos sociales.
Básicamente, las críticas neuroanatómicas a la teoría polivagal argumentan puntos que no se exponen en la teoría. Desde el enunciado inicial de la teoría, se ha hecho hincapié en los atributos únicos de la regulación neuronal del sistema nervioso autónomo de los mamíferos.
Este enfoque se destaca en el título de la publicación inicial – Orientarse en un mundo defensivo: Modificaciones en los mamíferos de nuestra herencia evolutiva. Una teoría polivagal. Funcionalmente, la teoría explicitó la capacidad única de los mamíferos de utilizar el comportamiento social a través del vago ventral y el sistema de conexión social para calmar los circuitos autónomos que apoyan la defensa (por ejemplo, el papel del sistema nervioso simpático en los comportamientos defensivos de lucha-huida y el papel del vago dorsal en la inmovilización defensiva). La tabla siguiente ofrece un resumen sucinto de las diferencias entre lo que afirma la teoría y lo que han dado por supuesto las críticas.
| Lo que afirma la teoría polivagal (Porges, 1995, 1998, 2007b) | Supuestos inexactos indicados (por ejemplo, Grossman y Taylor, 2007; Monteiro y col., 2018; Campbell y col., 2005). | |
| 1 | Se centra únicamente en las características únicas del sistema nervioso autónomo de los mamíferos que los distinguen de los reptiles. | Se centran en las características comunes del sistema nervioso autónomo en todas las especies de vertebrados. |
| 2 | Los mamíferos tienen una vía vagal mielinizada única que se origina únicamente en el núcleo vagal ventral (es decir, el núcleo ambiguo). | Las vías vagales mielinizadas son exclusivas de los mamíferos y las observaciones de fibras vagales mielinizadas procedentes del núcleo vagal dorsal en vertebrados distintos de los mamíferos refutan la teoría polivagal. |
| 3 | La arritmia sinusal respiratoria, definida como una interacción respiratoria-cardíaca exclusiva de los mamíferos, indexa la salida de las fibras cardioinhibitorias vagales ventrales que se originan únicamente en el núcleo vagal ventral (es decir, el núcleo ambiguo). | La arritmia sinusal respiratoria define las interacciones respiratorias y cardíacas observadas en todos los vertebrados que implican influencias vagales, incluidas las fibras cardioinhibitorias vagales originadas en el núcleo vagal dorsal. |
| 4 | Las fibras cardioinhibitorias vagales originadas en el núcleo vagal dorsal aparecen antes en la historia filogenética de los vertebrados que las fibras cardioinhibitorias vagales originadas en el núcleo ambiguo (véase más adelante). | No hay pruebas de que el núcleo vagal dorsal sea una fuente parasimpática del tronco encefálico evolutivamente más antigua que el núcleo ambiguo. |
El cuadro anterior resume las críticas a la teoría polivagal vinculadas a supuestos inexactos. Obsérvese que las críticas parecen reflejar una mala comprensión del alcance de la teoría. Las críticas se basan principalmente en las publicaciones de un científico comparativo consumado. Este científico y sus colegas han supuesto que la teoría polivagal puede ser falseada por las observaciones de las interacciones respiratorias y cardíacas y las fibras vagales mielinizadas en vertebrados que evolucionaron antes que los mamíferos. Nótese que la teoría polivagal, al centrarse en la transición desde los reptiles asociales hasta los mamíferos sociales, no guarda relación con esas críticas.
En consonancia con este malentendido, se ha criticado que no hay pruebas de que el núcleo vagal dorsal sea una fuente parasimpática del tronco encefálico evolutivamente más antigua que el núcleo ambiguo. Este punto de vista erróneo se basa en la suposición de que la presencia de neuronas vagales preganglionares ventrolaterales al núcleo vagal dorsal equivale a confirmar la existencia del núcleo ambiguo. La investigación neuroanatómica y neurofisiológica documenta una tendencia filogenética hacia una diferenciación en el área del tronco encefálico, de la que proceden las neuronas preganglionares vagales, en un núcleo vagal dorsal y un núcleo ambiguo discreto. No hay duda de que tanto las neuronas vagales cardíacas como las no cardíacas pueden encontrarse fuera del núcleo vagal dorsal en los vertebrados más primitivos. La filogenia del vago ilustra, a nivel neuroanatómico, la diferenciación de la columna eferente visceral del vago en un núcleo motor dorsal y un núcleo motor ventrolateral (es decir, el núcleo ambiguo), que se observa por primera vez en algunos reptiles. Por lo tanto, se podría suponer que la identificación neuroanatómica de un núcleo ambiguo discreto se limita a todos los mamíferos y a algunos reptiles. Esto no excluye la tendencia evolutiva de que las neuronas vagales preganglionares se encuentren ventrolaterales al núcleo vagal dorsal. Sin embargo, la función del núcleo ambiguo como fuente de las vías cardioinhibitorias vagales es exclusiva de los mamíferos.
Aunque los nuevos hallazgos en la literatura sobre la filogenia del sistema nervioso autónomo son interesantes y relevantes para los neuroanatomistas comparativos, estos hallazgos son relevantes para la teoría polivagal. Por ejemplo, la identificación de fibras vagales mielinizadas originadas en el núcleo vagal dorsal en otros vertebrados sugiere una serie de preguntas relacionadas con la función adaptativa y si existen fibras vagales mielinizadas originadas en el núcleo vagal dorsal en mamíferos que aún no han sido identificadas.
Utilizando la evolución y la maduración del desarrollo para trazar la jerarquía de los circuitos autónomos, la teoría polivagal propone un mapa vívido de la secuencia de respuesta a través de la cual progresan los mamíferos cuando se encuentran bajo amenaza, incluyendo enfermedades y lesiones. Esta secuencia que sigue a la evolución a la inversa, o disolución, como la describió sucintamente John Hughlyns Jackson (1884), refleja los relatos que experimentan con frecuencia los supervivientes de un trauma. Básicamente, las críticas no son críticas a la teoría polivagal. Las críticas no están relacionadas con la comprobación de los constructos documentados incluidos en la teoría: disolución, freno vagal, neurocepción o sistema de conexión social. Las críticas tampoco están relacionadas con el mayor conocimiento de los mecanismos implicados en el reclutamiento o el control del circuito vagal dorsal en los mamíferos. Más bien, las críticas, en el mejor de los casos, son tangenciales a la teoría y, en el peor, son representaciones inexactas que conducen a malentendidos de la teoría.
Otra serie de críticas tienen su origen en los trabajos de estos mismos científicos y se centran en el papel que desempeña la arritmia sinusal respiratoria dentro de la teoría polivagal. La tabla siguiente resume los puntos principales. En concreto, sostienen que existe una desconexión entre la arritmia sinusal respiratoria y el tono vagal cardíaco. La investigación ha documentado (véase Lewis y col., 2012) que la metodología de Grossman es defectuosa en la estimación del tono vagal ventral y es directamente la causa de su inferencia errónea con respecto a la inexactitud de la cuantificación de la arritmia sinusal respiratoria como un índice válido del tono vagal ventral. La investigación ha documentado (véase Lewis y col., 2012) la precisión y exactitud de la metodología de Porges-Bohrer y la contrasta con los métodos inadecuados utilizados por Grossman. Los defectos del método de Grossman se conocen y se documentan desde hace unos 25 años (Byrne y Porges, 1996; Lewis y col., 2012).
| Qué afirma la teoría polivagal | Supuestos inexactos indicados (por ejemplo, Grossman y Taylor, 2007) | |
| 1. | La arritmia sinusal respiratoria refleja con precisión el tono vagal cardíaco a través de las fibras vagales cardioinhibitorias mielinizadas originadas en el núcleo ambiguo. | Desconexión entre la arritmia sinusal respiratoria y el tono vagal cardíaco. |
| 2. | La teoría polivagal no se basa en que la arritmia sinusal respiratoria sea un índice preciso del tono vagal cardíaco. | La teoría polivagal se basa en que la arritmia sinusal respiratoria es exclusiva de los mamíferos. |
| 3. | La arritmia sinusal respiratoria es exclusiva de los mamíferos y refleja únicamente la influencia cardioinhibitoria de las fibras vagales procedentes del núcleo ambiguo. | La teoría polivagal puede refutarse observando la arritmia sinusal respiratoria en otros vertebrados determinada por fibras cardioinhibitorias originadas en núcleos vagales ventrales o dorsales. |
| 4. | La desconexión entre la arritmia sinusal respiratoria y el tono vagal cardíaco depende de la métrica utilizada para cuantificar la arritmia sinusal respiratoria. | La teoría polivagal puede refutarse documentando una desconexión entre la arritmia sinusal respiratoria y el tono vagal cardíaco, independientemente de la metodología específica utilizada para cuantificar la arritmia sinusal respiratoria. |
| 5. | Las hipótesis relacionadas con la teoría polivagal sólo pueden comprobarse con la arritmia sinusal respiratoria si la métrica proporciona un índice preciso del tono vagal cardiaco originado en el núcleo ambiguo. | Las hipótesis polivagales pueden comprobarse con cualquier medida de arritmia sinusal respiratoria. |


